СТРУКТУРА ДРЕВЕСНОЙ СИНУЗИИ ПИХТО-ЕЛЬНИКОВ КРУПНОПАПОРОТНИКОВЫХ В ВЕРХОВЬЯХ РЕКИ ПЕЧОРА (ПЕЧОРО-ИЛЫЧСКИЙ ЗАПОВЕДНИК)
Алейников А. А. (ЦЭПЛ РАН, г.Москва, РФ)
Лазников А.А., Ефименко А.С. (БГИТА, Брянск, РФ)
The structure of large-fern forests that
grow in spontaneously regime for a long time was investigated. Features of
plant composition and touchwood affection, core forest estimation
characteristics, age and ontogenesis structure were examined
for trees compartments.
Печоро-Илычский заповедник - наиболее крупный массив среднетаежных лесов в Европейской части России. Наибольший научный интерес для познания структуры и строения спонтанно развивающихся лесов представляют лесные массивы в труднодоступных верховьях реки Печора. С 2007 года ЦЭПЛ РАН проводит комплексные исследования биогеоценотического покрова бассейна реки Б. Порожняя (правый приток реки Печора) как модельного массива малонарушенных среднетаежных лесов.
В результате предварительных исследований на модельном полигоне выделены и описаны основные группы типов леса (Смирнова и др., 2007). В последующем была уточнена и дополнена классификация растительности (Смирнов, Браславская, 2010; Смирнов, 2010) и особенности возрастной структуры древостоев (Абатуров и др., 2010). Дешифрирование космоснимков показало, что крупнопапоротниковые леса достаточно широко распространены в модельном массиве. Первые исследования этих сообществ, проведенные в 2005 году (Смирнова и др., 2006, 2007), установили их пирогенный характер. Однако, последующие детальные исследования почв (Бовкунов и др., 2010), не подтвердили пирогенное происхождение сообществ (полное отсутствие углей в почвенных профилях), что может быть объяснено давностью пожара и последующим смывом углей. При внимательном рассмотрении мезорельефа, оказалось, что большая часть уже отмерших деревьев (в возрасте 250-300 лет) произрастала на элементах ветровально-почвенных комплексов, образованных предыдущими поколениями деревьев. Таким образом, можно предположить, что это сообщество уже несколько столетий развивается в спонтанном режиме без внешних воздействий.
Для изучения структуры и состава древесной синузии пихто-ельника крупнопапоротникового заложена постоянная пробная площадь размером 1 га (50х200 м). Она была разделена на 4 части по 50х50 м. На всей пробной площади проведен сплошной перечет древостоя, в который попали все деревьев с D1.3м>2 см. Для всех деревьев перечетного диаметра (n=1067) по последним общепринятым методикам (Методы…, 2002; Методические…, 2010) определялись: онтогенетическое состояние, календарный возраст, жизненность, высота и диаметр. Для определения абсолютного возраста произведено сплошное кернение деревьев перечетного диаметра на площади 50*100 м.
На территории модельного массива пихто-ельники крупнопапоротниковые занимают хорошо дренированные участки водоразделов и верхних частей склонов. Дешифрирование космоснимков (Iconos, 1 м/пикс.) показало, что площадь сообщества, в котором была заложена пробная площадь, составляет около 35 га. Древесный ярус сложен несколькими видами: елью сибирской (Picea obovata), пихтой сибирской (Abies sibirica), кедром сибирским (Pinus subirica) и березой пушистой (Betula pubesсens). Единично встречается древовидная рябина обыкновенная (Sorbus aucuparia). По сумме площадей поперечных сечений в древесном ярусе преобладает пихта и ель, значительно меньше кедра и березы. Состав древостоя: 4,9Пх3,0Е2,0К0,1Б. По числу стволов также доминируют пихта и ель, а вот березы в два раза больше, чем кедра. Преобладание конкурентных темнохвойных видов в составе древостоя и полное отсутствие реактивных светлохвойных (сосны и лиственницы) еще раз подтверждает, что последнее массовое нарушение этого участка было очень давно.
Древесный ярус сообщества очень неравномерен как по сомкнутости, так и по структуре. Распределение деревьев групповое, сомкнутость древесного полога небольшая (0,3-0,4). В целом, древостой характеризуется вертикальной сомкнутостью и отсутствием четких ярусов, что значительно затрудняет использование стандартных методик (Анучин, 1982; Лесотаксационный…, 1984) для определения таксационных показателей. Значительная внутри- и межвидовая конкуренция, свойственная разновозрастным лесам приводит к сильному варьированию основных таксационных характеристик.
Таблица 1 - Характеристика древесной синузии пихто-ельника крупнопапоротникового
|
Породы |
Число живых деревьев, шт./га |
Площадь поперечного сечения, м2 |
Число классов возраста* |
Макс. возраст, лет** |
Макс. диаметр, см |
Макс. высота, м |
|
Пихта |
508 |
15,42 |
6 |
<200 |
51,6 |
28,7 |
|
Ель |
351 |
9,50 |
7 |
<250 |
56,1 |
30,0 |
|
Кедр |
16 |
6,20 |
2 |
<450 |
97,5 |
28,0 |
|
Береза |
37 |
0,37 |
- |
- |
34,9 |
18,7 |
|
Итого |
912 |
31,49 |
|
|||
Примечание: *Класс возраста - 40 лет
**Более точно определить возраст невозможно из-за стволовой гнили.
Определение абсолютного возраста показало, что по классификации С. А. Дыренкова (1984), ценопопуляции пихты и ели относятся к абсолютно-разновозрастным и представлены 6-7 классами возраста (1 класс -40 лет), что означает одновременное присутствие в древостое нескольких возрастных поколений. Возобновление этих видов идет постоянно. Кедр представлен 2 классами возраста (двумя поколениями), возобновление его очень затруднено.
Анализ распределения доминантов древесного яруса (пихты и ели) по ступеням толщины (2 см) показал, что у обоих видов максимум смещен влево и с увеличением диаметра число деревьев постепенно уменьшается. Схожая закономерность наблюдается и в распределение деревьев по двухметровым ступеням высоты.
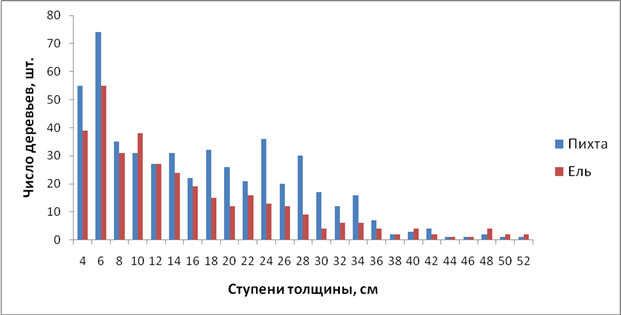
Рисунок 1 - Распределение стволов пихты и ели по ступеням толщины
Абсолютную разноразмерность по диаметру и высоте доминирующих видов древостоя подтверждают и статистические показатели, приведенные в табл. 2. Коэффициенты асимметрии (S), отличающиеся от нуля подтверждают, что распределение не симметрично и максимум деревьев смещен влево. Различия в этом коэффициенте показывают, что распределение деревьев пихты изменяется более плавно, чем ели.
Таблица 2 - Статистические показатели распределения по диаметру и высоте
|
Порода |
Статистические показатели распределения |
|||||||
|
по диаметру |
по высоте |
|||||||
|
M±m |
δ |
Е |
S |
M±m |
δ |
E |
S |
|
|
Пихта |
16,7±0,46 |
10,40 |
-0,33 |
0,63 |
11,23±0,30 |
6,85 |
-1,01 |
0,36 |
|
Ель |
15,09±0,58 |
10,83 |
1,69 |
1,36 |
9,34±0,36 |
6,79 |
0,08 |
0,97 |
Связь между высотой и диаметром у пихты и ели довольно высокая (R > 0.8). А связь между этими таксационными показателями и возрастом отсутствует (или очень слабая). Такая особенность характерная для всех разновозрастных лесов (Дыренков, 1984; Бобкова, 2006).
Демографический анализ показал, что нормальные онтогенетические спектры ценопопуляций с высокой плотностью имеют оба доминирующих вида: пихта и ель. Они представлены в древостое всеми онтогенетическими состояниями (от v1 до g3), причем среди них преобладают молодые генеративные особи (26% и 29% соответственно). Ценопопуляция кедра имеет регрессивный спектр с малой плотностью особей и представлена только генеративными растениями, среди которых доминируют старые генеративные (73%). Ценопопуляция березы имеет спектр близкий к нормальному, но отличается малой плотностью особей: единично встречаются виргинильные и молодые генеративные растения березы.
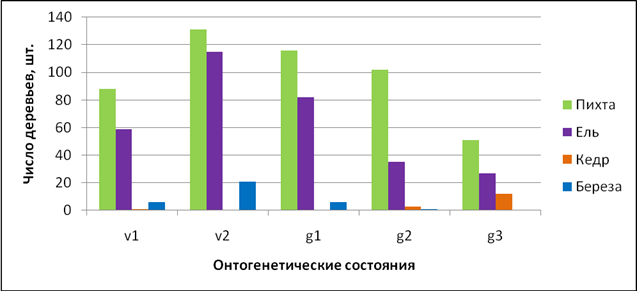
Рисунок 2 – Онтогенетический спектр древесной синузии пихто-ельника крупнопапоротникового
Исследование санитарного состояния древостоя показало, что в древесном ярусе велика доля деревьев, пораженных фитопатогенными грибами (Atmillaria mellea, Fomitopsis annosa, Onnia triqueter, Phellinus chrysoloma и др.) Для анализа распространения гнилей исследованы 191 пихта и 142 ели. Оказалось, что, несмотря на общие патогенные виды, пораженность пихты значительно превосходит пораженность ели (47% у пихты и 8% у ели). Распределение пораженных деревьев ели и пихты по ступеням толщины показано на рис. 3.
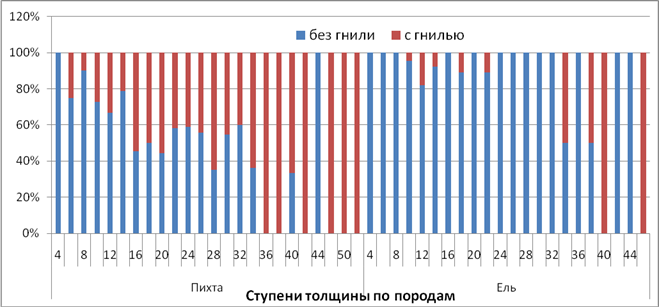
Рисунок 3 – Соотношение деревьев пихты и ели с гнилью и без гнили по ступеням толщины
Значительная пораженность деревьев комлевыми и стволовыми гнилями приводит к тому, что большая часть деревьев обламывается на разной высоте, а не вываливается. Отсутствие педотурбаций препятствует увеличению биоразнообразия в целом и приживанию подроста деревьев.
Таким образом, детальные исследования выявили особенности состава и строения древесного яруса пихто-ельника крупнопапоротникового. Подтверждена сложная структура древостоя: абсолютная разновозрастность и разноразмерность. В ближайшем будущем, при отсутствии внешних воздействий, в древостое по-прежнему будут преобладать пихта и ель. А вот кедр, скорее всего, будет вытеснен из этого сообщества (Алейников и др., 2011; Ефименко и др., 2011). Рекогносцировочные исследования в заповедниках Урала и восточной части равнинной территории Коми выявили отсутствие молодых поколений в ценопопуляциях кедра на дренированных (автоморфных) позициях. Таким образом, проблема деградации нормальной возрастной структуры ценопопуляций кедра в природных лесах на западной границе ареала приобретает важное значение и требует разносторонних дополнительных исследований биогеоценотического уровня.
Работа выполнена при финансовой поддержке фонда РФФИ (проект № 10-04-00355) и Программы фундаментальных исследований Президиума РАН «Научные основы сохранения биоразнообразия России».
Литература
1. Абатуров А.В, Браславская Т. Ю, Кулешов А.П., Алейников А. А. Некоторые особенности возрастной структуры темнохвойных лесов Печоро-Илычского заповедника // Труды Печоро-Илычского заповедника. Сыктывкар, 2010. Вып. 16. С. 5-10.
2. Алейников А. А., Лазников А. А., Ефименко А. С. Онтогенетическая структура ценопопуляции кедра в крупнопапоротниковых лесах бассейна реки Б. Порожняя / Биология – наука XXI века: Тез. 15 Межд. Пущинской школы-конф. молодых ученых. – Пущино, 2011. - С. 247-248.
3. Анучин Н. Д., Лесная таксация. М., 1982. – 552 с. 1982
4. Бобкова К. С., Галенко Э. П., Загирова С. В., Патов А. И., Сенькина С. Н., Машика А. В., 2005 Коренные еловые леса предгорного ландшафтного района Печоро-Илычского заповедника / Труды Печоро-Илычского заповедника. Вып. 14. 2005. – с. 19-25.
5. Бобкова, 2006 Коренные еловые леса Севера: биоразнообразие, структура, функции. – Спб., 2006. – 337 с.
6. Бовкунов А.Д., Семиколенных А. А., Алейников А. А., Ухтомский В. Г. Основные типы почв темнохвойных лесов нижнего участка бассейна реки Большая Порожняя (приток р. Печора) / Труды Печоро-Илычского заповедника. Сыктывкар, 2010. Вып. 16. С. 23-31.
7. Дыренков С. А. Структура и динамика таежных ельников. Л., 1984. – 175 с., 1984
8. Ефименко А. С., Лазников А. А., Алейников А. А. Особенности возобновления кедра сибирского в различных типах леса бассейна реки Б. Порожняя (Печоро-Илычский заповедник) / Ломоносов -2011: XVIII Межд. конф. Студентов, аспирантов и молод. Ученых; Секция «Биология»; Москва, МГУ им. М. В. Ломоносова: Тезисы докладов. М., 2011. – с. 319.
9. Лесотаксационный справочник для Северо-Востока европейской части СССР / Отв. Сост. Г.С.Войнов. Архангельск, 1986. 357 с.
10. Методические подходы к экологической оценке лесного покрова в бассейне малой реки / Отв. ред. Л.Б. Заугольнова, Т.Ю. Браславская. - М.: 2010. 383 с.
11. Методы изучения лесных сообществ. –Спб., 2002. – 240 с.
12. Смирнов Н. С. Основные типы темнохвойных лесов бассейна р. Б. Порожняя (Печоро-Илычский заповедник) // Известия Самарского НЦ РАН. 2010. т.12 (33). №1 (3). С. 892 – 896.
13. Смирнова О.В., Бобровский М.В., Ханина Л.Г., Смирнов В.Э. Биоразнообразие и сукцессионный статус темнохвойных лесов Шежемпечерского и Большепорожского ботанико-географического районов Печоро-Илычского заповедника // Труды Печоро-Илычского заповедника. Вып. 15. 2007. Стр. 28-47.
14. Смирнова О.В., Бобровский М.В., Ханина Л.Г., Смирнов В.Э. Биоразнообразие и сукцессионный статус старовозрастных темнохвойных лесов Европейской России // Успехи современной биологии. 2006. т. 126, №1. Стр. 27-49.
15. Смирнова О.В., Торопова Н. А., Луговая Д. Л., Алейников А. А. Методология исследования популяционной организации и сукцессионной динамики лесных экосистем (биогеоценозов) / Методические подходы к экологической оценке лесного покрова в бассейне малой реки / Л.Б. Заугольнова, Т.Ю. Браславская (отв. ред.). М.: 2010. С. 20 - 40.