О ПРОДОЛЖИТЕЛЬНОСТИ ЖИЗНИ ХВОИ КАВКАЗСКОЙ ПИХТЫ
Придня М.В., Шолохов А.Г., Савков А.А.
(ФГУ «НИИгорлесэкол», г. Сочи, РФ)
By help of statistical comparison is shown that the growth of foliage of full-leaves phenotype have trend to exceed its of alternative hole-leaves phenotype of Caucasian fir.
The growth of foliage on lateral branch exceeds its on basal stem, that connected with their good exposition to light.
Продолжительность жизни хвои существенно различается в пределах вида, а также среди разных видов и таксонов более высокого ранга. Не только в пределах вида или его отдельной популяции этот показатель существенно различается даже в пределах отдельной семьи кавказской пихты, нами наблюдались различия продолжительности жизни хвои на порядок величин (4-40 лет). Поэтому, по-видимому, вполне оправданно изучение специфичности проявления данного показателя, как для видов и его подразделений, так и для надвидовых таксонов, в качестве индикаторов оценки жизненности растений, в том числе и зависящей от окружающей среды (биотической и абиотической) (Ярмишко, 2001).
Нами выполнен сравнительный анализ продолжительности жизни хвои разных фенотипов кавказской пихты в одной семье (Придня, 1975, 1977). В Колхидской подпровинции в Причерноморской провинции Северного Кавказа, в бассейне р. Шахе в Западном отделе КГПБЗ. При этом сравнивались отдельно показатели продолжительности жизни осевого побега цельнолистного и выямчатолистного фенотипов, а также бокового побега этих же фенотипов. После вычисления статистических показателей (таблица) были вычислены критерии достоверности различия t (Стьюдента) по 16 парам одновозрастных рядов сравнения цельнолистного фенотипа и 11 парам выямчатолистного. При этом возраст растений колебался при сравнении хвои осевого побега от 14 до 26 лет, а при сравнении бокового побега – от 14 – до 30 лет. Обусловлено это тем, что в данной семье произведен полный учет всех особей от 1 до 62 лет, в анализ включен наличный резерв особей в семье по этим возрастным группам. Этим же обусловлено и число особей по возрастным генерациям.
Получены следующие результаты. Прежде всего, по всему спектру возрастной структуры семьи, сложенной более 1200-ми особями, с указанными возрастными генерациями возраст хвои колебался от 1 до 40 лет. В таблице приведены наиболее часто встречающиеся продолжительности жизни (возраста) хвои в наиболее массовых генерациях. Соотношения средних возрастов хвои по фенотипам и по принадлежности хвои (боковой побег или осевой) приведены на рисунках 1 и 2, полученных в качестве примеров из другой семьи этого вида, на значительном удалении и возвышении от первой.
Из этих материалов вытекают следующие результаты.
1. Из 16 возрастных генераций хвоя на осевом побеге цельнолистного фенотипа имеет достоверно большую продолжительность жизни в 6 случаях, т. е. в 37,5% случаев, чем у выямчатолистного. В 8 случаях (50%) возраст хвои цельнолистного выше такового выямчатолисного, хотя различия не достоверны, т. е. можно говорить о тенденции превышения продолжительности жизни хвои этого фенотипа. В двух случаях (12,5%) наблюдается тенденция превышения возраста выямчатолистного фенотипа над таковым цельнолистного (различия не достоверны).
2. Из 11 генераций в 4 (36,4%) случаях хвоя на боковом побеге цельнолисного фенотипа превосходит по продолжительности жизни таковую у выямчатолистного фенотипа. В 5 случаях (45,4%) хвоя цельнолистного фенотипа превосходит по этому показателю её у выямчатолистного (не достоверно), т. е. имеется тенденция превышения. А в двух парах (18,2%) также выямчатолистный фенотип имеет тенденцию превышения по этому показателю над цельнолистным (различия не достоверны). Как видим, продолжительность жизни хвои цельнолистного фенотипа имеет тенденцию превышать её у выямчатолистного как на осевом, так и на боковом побегах.
3. Из приведенных материалов (графики и таблица) видно, что продолжительность жизни хвои на боковых побегах превышает таковую на осевых, что вполне объяснимо более выгодным положением первой по отношению ко второй в смысле доступности солнечного света. Показатель продолжительности жизни хвои может служить не только индикатором состояния семьи или популяции (вида), но даже органов в пределах особи; использование его требует разработки специальных статистических подходов, наши данные могут быть использованы для этой цели. Хотя абсолютные показатели различия варьирует в пределах 1-4 лет, эти различия в масштабах многочисленных популяций и семьях имеют существенное значение при решении ими проблемы выживания при динамично меняющихся условиях биотической и абиотической среды.
Таблица - Сравнение возраста хвои выямчатолистного и цельнолистного фенотипов
Осевой побег |
|||||||
Фенотипы |
Возрастдерева, лет |
Возрастхвои, лет |
N |
м |
§ |
р |
Тф /Tst |
В |
14 |
6,7 |
7 |
0,36 |
0,95 |
5,4 |
4,6 2,45 |
|
Ц |
14 |
8,6 |
7 |
0,61 |
1,62 |
7,1 |
|
|
В |
15 |
6,8 |
23 |
0,31 |
1,47 |
4,5 |
2,83 2,07 |
|
Ц |
15 |
8,3 |
23 |
0,3 |
1,45 |
3,7 |
|
|
В |
20 |
8,1 |
55 |
0,22 |
1,66 |
2,8 |
4,38 2 |
|
Ц |
20 |
9,5 |
55 |
0,24 |
1,78 |
2,5 |
|
|
В |
21 |
8,3 |
21 |
0,45 |
2,08 |
5,4 |
3,29 2,09 |
|
Ц |
21 |
10,2 |
21 |
0,47 |
2,16 |
4,5 |
|
|
В |
24 |
7,5 |
15 |
0,32 |
1,25 |
4,3 |
3,37 2,14 |
|
Ц |
24 |
9,5 |
15 |
0,58 |
2,26 |
6,1 |
|
|
В |
25 |
8,7 |
48 |
0,28 |
1,95 |
3,24 |
3,56 2,01 |
|
Ц |
25 |
10,1 |
48 |
0,34 |
2,35 |
3,4 |
|
Боковой побег |
|||||||
|
В |
14 |
7,7 |
7 |
0,47 |
1,25 |
6,1 |
3,27 2,45 |
|
Ц |
14 |
9,3 |
7 |
0,18 |
0,49 |
2 |
|
|
В |
15 |
8 |
23 |
0,36 |
1,72 |
4,5 |
4,01 2,07 |
|
Ц |
15 |
10,1 |
23 |
0,31 |
1,47 |
3 |
|
|
В |
25 |
11,5 |
48 |
0,44 |
3,02 |
3,79 |
3,8 2,01 |
|
Ц |
25 |
13,7 |
48 |
0,36 |
2,52 |
2,64 |
|
|
В |
26 |
11,1 |
11 |
1,1 |
3,65 |
9,91 |
2,6 2,23 |
|
Ц |
26 |
13,8 |
11 |
0,83 |
2,75 |
6 |
|
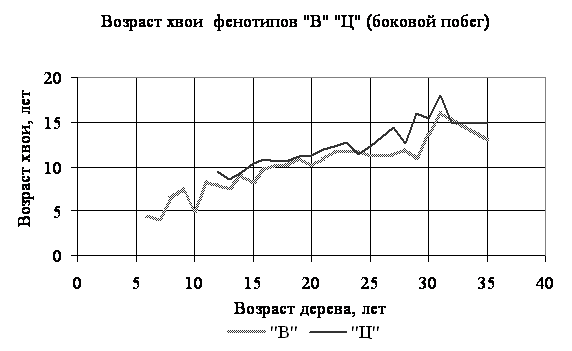
Рисунок 1 - Соотношения средних возрастов хвои по фенотипам от принадлежности хвои (боковой побег)
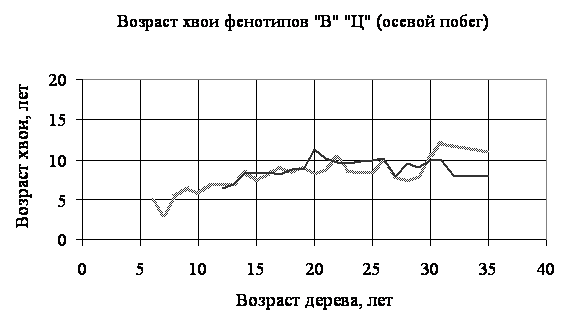
Рисунок 2 - Соотношения средних возрастов хвои по фенотипам от принадлежности хвои (осевой побег)